
|
Science » » Pierwsze błony komórkowe [2] Złapanie życia w ryzyPowyżej wspomniana publikacja, moim zdaniem, jest równie wartościowa i sensacyjna jak pierwsze eksperymenty Stanleya-Millera (opisywany w pierwszym artykule). Naukowcy elegancko udowodnili, że formujące się pęcherzyki mogą zamykać w swoim wnętrzu gotowe RNA oraz fragmenty montmorylonitu, na których wciąż tworzą się nowe cząsteczki kwasu rybonukleinowego. Twory takie na dodatek mają zdolność do biernego, podstawowego transportu [7] — zaobserwowano z czasem przenikanie barwnika do wnętrz tych swoistych „pre-komórek". P. Monnard, współpracujący z D.Deamerem, wykazał, że pęcherzyki formowane z kwasu i alkoholu dodekanowego mogą uwięzić mieszaninę cząsteczek DNA o długości około 600 par zasad (par nukleotydów) [2, 3]. Spójrz na poniższy rycina. 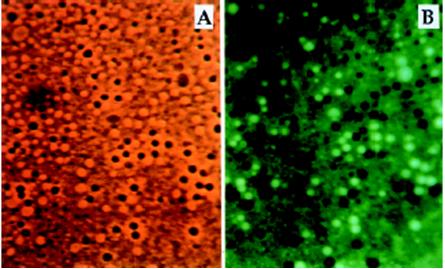 Rycina 6. Uwięzione DNA w pęcherzykach zbudowanych z kwasu i alkoholu dodekanowego. Pęcherzyki z DNA w panelu (A) są czarne, pod wpływem światła UV emitują intensywnie zielone światło. [3] RNA uwięzione w takim pęcherzyku miało większą szansę na stworzenie funkcjonalnego systemu biochemicznego, niż pływając swobodnie w roztworze. Zwiększa się w tym wypadku możliwość wspólnych interakcji i nabycia nowych funkcji. Zwiększa się stopień uporządkowania całego układu. Co więcej, aktywowane nukleotydy mogły dyfundować w obydwie strony poprzez błonę, dostarczając budulca do nowych łańcuchów RNA. Inną drogą nukleotydów do wnętrza pre-komórki były naturalnie występujące, krótkotrwale tworzące się przerwy w dwuwarstwie. Zagadnienie transportu stanowi poważny problem, ograniczając możliwości pierwszych systemów metabolicznych. Zakładamy raczej, że energia chemiczna była głównym źródłem zasilania w świecie prebiotycznym. A co z fotosyntezą? Czy ostatnie doniesienia o jej śladach 3,7 miliarda lat temu nie dają do myślenia? Aby wykorzystać energię słoneczną, fotony muszą być najpierw zaabsorbowane przez substancje pigmentowe i przekształcone w niestabilne formy energii chemicznej. Oczywiście systemy porównywalne do dzisiejszych komórek były jeszcze nieobecne, ale istniały wspomniane wcześniej związki z grupy PAH, które mogły zostać inkorporowane w zamknięte struktury dwuwarstwowe. Dotychczas pojawiło się kilka sygnałów o właściwościach PAH jako pigmentu [3]. Badania trwają. Dzisiejszy, wydajny, nawet bierny (zgodnie z gradientem stężenia przenoszonej substancji) transport oparty jest na białkach przenośnikowych. (Rycina 7). Świat komórkowy dysponuje również fascynującym i wyrafinowanym systemem transportu aktywnego (wbrew gradientowi stężęń — Rycina 8) na wiele sposobów regulowanym. 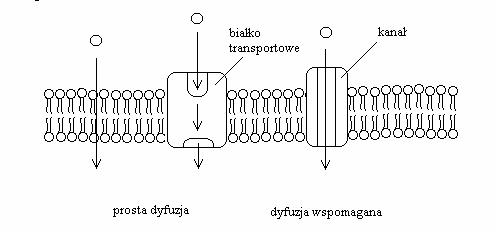 Rycina 7. Rodzaje transportu biernego współczesnych błon. Transport zachodzi zawsze zgodnie z gradientem stężeń przenoszonej cząsteczki - jeśli substancji jest więcej na zewnątrz, będzie ona transportowana do wnętrza, aby wyrównać stężenia. 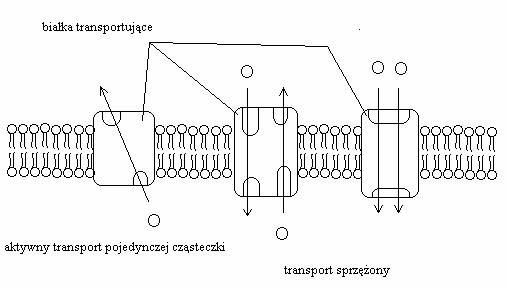 Rycina 8. Podstawowe rodzaje transportu aktywnego współczesnych błon. Transport zachodzi niezależnie od gradientu stężeń przenoszonych cząsteczek, ale wymaga dostarczenia energii. Innymi formami transportu aktywnego są również tutaj nie opisywane endo- i egzocytoza. Możemy spodziewać się więc, że wysoka efektywność przenoszenia rozpoczęła się dopiero po pojawieniu się pierwszych białek, które mogły być „złapane" w błonie. Nie zaobserwowano RNA tworzącego twory transbłonowe. Jak to się stało — jest dla nas na razie nierozwiązaną zagadką, ale trzymajmy kciuki za biologów symulujących powstanie życia... Czas na dzieci. Czas na lepszą ewolucjęWreszcie dotarliśmy do kwestii przekazania materiału genetycznego. Wrócimy tutaj do wcześniej wspominanego artykułu trójki naukowców. Postawili oni następującą tezę: prymitywne rozmnażanie najprawdopodobniej polegało na prostym podziale pęcherzyka razem z materiałem zawartym w środku. Jeśli wewnątrz pęcherzyka wykształci się pewien jednolity system, podzielenie go na pół da nam dwa pęcherzyki z systemami zdolnymi do niezależnych zmian i niezależnej ewolucji. Jednak aby utrzymać ten podział takie pra-komórki powinny rosnąć. Okazało się, że pęcherzyki rosną pod wpływem zwiększania się ilości miceli w roztworze. Czyli najpierw powstają micele, które przekształcają się w nowe dwuwarstwowe pęcherzyki, po czym po pewnym wysyceniu ogranicza się powstawanie nowych pęcherzyków a zwiększająca się wciąż ilość miceli. Cześć składników miceli — w na razie szczegółowo nie poznany sposób — łączy się z pęcherzykami zwiększając ich objętość. Proces ten dotyczy 90% puli pęcherzyków w roztworze [7]. Jak zachodzi podział? Otóż bynajmniej nie poprzez reakcje biochemiczne. Naukowcy przepuścili podrośnięte pęcherzyki przez porowaty materiał (o porach rozmiarów 100 nm) i okazało się, że podzieliły się ona na pół z bardzo małą stratą zawartego materiału. Następnie, aby potwierdzić produktywność tego systemu, przeplatali cykle wzrostu (dodając micele) oraz podziału (przepychając przez wspomnianą „gąbkę"). Całe doświadczenie dowiodło, że wzrost pęcherzyków jest możliwy dzięki naturalnemu wzrostowi stężenia miceli, zaś podział może zaistnieć dzięki prostym procesom fizykochemicznym [7]. P. Monnard i D. Deamer poszli o krok dalej — zaproponowali minimalny model komórki oparty na katalitycznym i strukturalnym RNA zobrazowany na Rysunku 9. 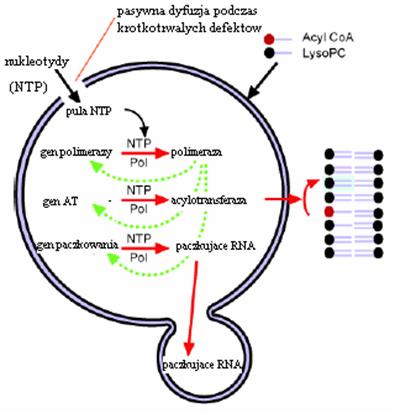 Rycina 9. Minimalny model komórki oparty na katalitycznym i strukturalnym RNA. Opis w tekście. [9] Mimo iż teoria fragmentów RNA o właściwościach polimerazy RNA (produkowania kopii RNA; zob. poprzednie artykuły) jest niekompletna, to jest całkiem prawdopodobna i rozwijana [8]. Naukowcy twierdzą, że niezbędne byłyby trzy systemy [3]. Jeden zawierałby gen polimerazy, na podstawie którego polimeraza produkowałaby swoje bliźniacze kopie. Enzym ten produkowałby również na podstawie innego genu acylotransferazę, która aktywnie brałaby udział we wzroście błony komórkowej. Wreszcie namnażałby cząsteczki RNA odpowiedzialne za "pączkowanie". Model nie ma na razie poparcia doświadczalnego. Jakimkolwiek torem podążał rozwój komórek — dzisiejsze efekty są imponujące. I pomyśleć, że około czterech, może i więcej miliardów lat temu wystarczyła iskierka. Resztę „roboty" wykonała ewolucja. Ufam, że pomogłem Czytelnikom Racjonalisty nieco przybliżyć obecny stan wiedzy o początkach życia na Ziemi. Tutaj na razie kończymy naszą wędrówkę, mimo że na horyzoncie widać wiele terenów do eksploracji. Może kiedyś zaproszę jeszcze na podobną eskapadę w nieporównywalnie bardziej skomplikowany i równie fascynujący świat współczesnych komórek i pra-organizmów. Dzisiejsza nauka potrafi przyprawić nas o zachwyt i coraz częściej zawroty głowy. Cieszę się ogromnie, że jestem świadkiem tak rewolucyjnych odkryć jak zeszłoroczny sukces Hanczyca, Fujikawy i Szostaka. Kilka tygodni temu koreańscy naukowcy, adaptując komórki macierzyste do hodowli, dokonali pierwszego zdecydowanego kroku w kierunku inżynierii tkankowej. Fizycy są coraz bliżej unifikacji, zwieńczenia ludzkiej nauki. Czekamy w napięciu na sygnały życia na Marsie. Mam nadzieję, że udało mi się chociaż trochę zarazić Czytelników moim entuzjazmem i zachęcić ich do własnych poszukiwań i rozmyślań nad początkami życia.
Literatura: Rysunki 2, 4, 5, 6 i 9 przerysowane, przetłumaczone i zamieszczone w artykule za zgodą dra Davida Deamera.
« (Published: 03-03-2004 Last change: 07-04-2004)
page 3272 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||



 Ukończył biologię molekularną na Uniwersytecie Adama Mickiewicza w Poznaniu. Pracował jako Research Specialist in Health Science w Department of Anatomy and Cell Biology na University of Illinois w Chicago. Obecnie pracuje jako Associate Cell Biologist / Histologist w Abbott Laboratories (Illinois). Specjalizuje się w ekspresji białek 'od zera', hodowlach linii komórkowych, symulacji in vitro procesów zachodzących w komórkach. Jego pasją jest teoria ewolucji, w szczególności ewolucja systemów biochemicznych i pochodzenie życia we Wszechświecie.
Ukończył biologię molekularną na Uniwersytecie Adama Mickiewicza w Poznaniu. Pracował jako Research Specialist in Health Science w Department of Anatomy and Cell Biology na University of Illinois w Chicago. Obecnie pracuje jako Associate Cell Biologist / Histologist w Abbott Laboratories (Illinois). Specjalizuje się w ekspresji białek 'od zera', hodowlach linii komórkowych, symulacji in vitro procesów zachodzących w komórkach. Jego pasją jest teoria ewolucji, w szczególności ewolucja systemów biochemicznych i pochodzenie życia we Wszechświecie.| [ Cooperation ] [ Advertise ] [ Map of the site ] [ F.A.Q. ] [ Store ] [ Sign up ] [ Contact ] The Rationalist © Copyright 2000-2018 (English section of Polish Racjonalista.pl) | ||